拟南芥苯丙氨酸解氨酶(PAL)基因的研究进展
本作品内容为拟南芥苯丙氨酸解氨酶(PAL)基因的研究进展,格式为 docx ,大小 42679 KB ,页数为 6页

('拟南芥苯丙氨酸解氨酶(PAL)基因的研究进展孙海燕;全雪丽;付爽;吴松权【摘要】苯丙氨酸解氨酶(PAL)是催化苯丙烷代谢途径第1步反应的限速酶,广泛地参与植物生长发育过程中的各种生理活动.本文概述了拟南芥PAL基因的分子生物学和生理学研究进展,主要包括PAL基因的结构、表达特性、调控机制及其参与的植物生理学的意义,为进一步阐明PAL基因的功能提供参考依据.【期刊名称】《延边大学农学学报》【年(卷),期】2016(038)001【总页数】5页(P88-92)【关键词】拟南芥;苯丙氨酸解氨酶;表达;生理作用【作者】孙海燕;全雪丽;付爽;吴松权【作者单位】延边大学农学院,吉林延吉133002;延边大学农学院,吉林延吉133002;延边大学农学院,吉林延吉133002;延边大学农学院,吉林延吉133002【正文语种】中文【中图分类】Q943苯丙烷代谢途径是陆生植物生长和发育所必需的,是植物长期适应自然环境的结果[1-2]。苯丙氨酸解氨酶(EC4.3.15,PAL)催化L-苯丙氨酸(L-Phe)的非氧化脱氨基作用,生成肉桂酸,是生物合成苯丙烷类天然产物的第1步[3-4],也是第1个被鉴定的植物“防御基因”[5]。也是苯丙烷类代谢途径中研究最多的酶[6]。肉桂酸是植物生长、发育和环境适应所必需的各种苯丙烷类物质的合成起始物[7]。苯丙烷类化合物是植物中大量酚类化合物的前体,包括木质素、黄酮类、异黄酮类、香豆素、芪类和水杨酸等,它们在维持植物结构、抵御紫外线、形成花青素和植保素、保持花粉活力、信号转导与交流等方面发挥着重要作用[1,5,8]。自1961年Koukol和Conn首次描述PAL以来,苯丙氨酸解氨酶得到了广泛的研究[9],它是控制苯丙烷途径生物合成去向的关键酶和限速酶[7,10]。拟南芥是一种十字花科植物,广泛用于植物遗传学、发育生物学和分子生物学的研究,已成为一种典型的模式植物,该植物具有个体小、生长周期快、形态特征简单、生命力强、基因组小、遗传操作简单等优点[11-12]。通常PAL基因在植物中编码小的基因家族[9],苯丙氨酸解氨酶的同工酶负责苯丙烷途径的不同分支途径[10],拟南芥有4个PAL同工酶,它们的表达受植物本身的生长发育调节和环境胁迫所诱导[1-3,7-10],本文主要介绍过去20多年来拟南芥植物4个PAL基因的分子生物学和生理学的研究进展,为进一步阐明PAL基因家族在拟南芥植物生长发育过程中所发挥的功能提供参考依据。Southern杂交[4]和基因组测序表明拟南芥基因组编码4个不同的PAL基因[2-3,7,9],分别是AtPAL1(AT2G37040)、AtPAL2(AT3G53260)、AtPAL3(AT5G04230),和AtPAL4(AT3G10340)[7],分别定位于拟南芥5条染色体中的3条染色体上,AtPAL1定位于染色体Ⅱ,AtPAL2和AtPAL4定位于染色体Ⅲ,AtPAL3定位于染色体Ⅴ的端粒附近[9]。AtPAL编码长度为2100bp左右,变化幅度为24~81bp。其中,AtPAL1编码长度为2178bp,AtPAL2编码长度为2154bp,AtPAL3编码长度为2097bp,AtPAL4编码长度为2124bp。DNA序列分析表明,PAL基因家族分为2个亚族,AtPAL1和AtPAL2属于1个亚族,AtPAL3和AtPAL4属于另一个亚族,这种2+2方式可能是复制的结果,因为AtPAL1和AtPAL2一致性达90.4%,AtPAL3和AtPAL4一致性达83.5%。2个亚族间所含内含子数目也不同,AtPAL1和AtPAL2中有1个内含子,AtPAL3和AtPAL4中有2个内含子,表明这2个亚族转录后加工的机制不同[9]。生物信息学分析表明,这4个AtPAL蛋白质在氨基酸水平上具有≥74%的一致性[9],所有AtPAL蛋白定位在细胞质中并缺乏目标多肽序列[2]。4个AtPAL的启动子在转录起始位点上游500bp处都存在所谓的PAL盒,AtPAL1、AtPAL2、AtPAL3和AtPAL4分别具有4、3、1和4个PAL盒[13]。AtPAL1启动子和AtPAL2启动子存在4个盒1序列(或L盒),而AtPAL3启动子存在1个;AtPAL1启动子存在3个盒2序列(或P盒),而AtPAL2启动子和AtPAL3启动子各自存在1个;AtPAL1启动子存在13个盒3(AC元件)序列,AtPAL2启动子存在9个,而AtPAL3启动子存在2个[5]。AtPAL4启动子中存在A盒、H盒和G盒,但缺乏AC元件[14]。综上所述,AtPAL1和AtPAL2不仅在结构上非常相似而且其启动子元件也非常相似。AtPAL原核表达分析表明,每1个AtPAL都具有独特的催化活性[9]。AtPAL1,2和4在低浓度(0.5~1μg)时对L-苯丙氨酸(L-Phe)具有催化活性,而AtPAL3在高浓度(≥50μg)时对L-Phe具有催化活性。4个酶最适pH值相近(8.4~9.2),但所需的最佳反应温度却不同,相对于其它3个酶(46~48℃),AtPAL3的最佳反应温度较低(31℃)。AtPAL1,2和4对L-苯丙氨酸具有相似的米氏常数(64~71),AtPAL2的催化效率最高。此外,AtPAL1,2和4对L-苯丙氨酸动力学曲线遵循典型的米氏方程。当L-酪氨酸(L-Tyr)为底物时,AtPAL1,2和4催化效率较低,而AtPAL3则没有催化活性,因此,L-Phe才是AtPAL真正的生理学底物。这和预期结果一致,因为双子叶植物PAL不能有效利用L-Tyr作为底物。顺式和反式肉桂酸对AtPAL催化活性有不同的影响,因为顺式肉桂酸是AtPAL1的竞争性抑制剂,而不是AtPAL2和AtPAL4的竞争性抑制剂,但是反式肉桂酸是AtPAL1,2和4的竞争性抑制剂[15]。AtPAL的表达具有组织特异性和时空表达差异性。4个AtPAL在种子、幼苗、根、花序茎、叶、花以及角果中都表达,一般情况下AtPAL3表达水平较低[14],且在幼苗及根中的表达水平最低[5],在植物发育过程中,AtPAL1和AtPAL2表达方式非常相似,尤其是在根和花序茎中[4-5,7]。AtPAL1在幼苗发育早期[4,16]和成熟组织中激活,并且大量表达在根和叶的维管束组织中,但是在根尖或顶端分生组织中不表达[4]。AtPAL1在花的萼片、花药和心皮中表达,但是在白色的无色素的花瓣中不表达[4],而AtPAL2在花瓣和花药中都不表达[7]。AtPAL的表达调控机制复杂,它的表达不仅受到植物本身发育水平的调控,还受到非生物和生物因素的诱导。AtPAL在拟南芥生长发育过程中的表达具有组织特异性,受发育的调控。AtPAL1在种植2d的拟南芥幼苗中的表达量最高,之后随着植物生长发育,表达量逐渐降低[4]。尽管AtPAL1的表达量比AtPAL2多,但它们在根、茎和叶中的表达方式却相同,不过AtPAL3只在生长10d的幼苗和成熟植物根中表达[5]。这应与基因结构有关,因为AtPAL1和AtPAL2启动子区序列具有高度的相似性,而AtPAL3启动子区序列与AtPPAL1和AtPAL2启动子区序列缺乏序列相似性[5],且AtPAL1和AtPAL2编码区的相似性大于它们与AtPAL3的相似性。在花序茎生长过程中(1~20cm),AtPAL1和AtPAL2的表达量持续增加;当花序茎逐渐生长到10cm之后,AtPAL4的表达趋于稳定;而AtPAL3的表达呈波动形式[14]。AtPAL2参与植株营养生长到生殖生长的整个过程中[7];花芽发育中,AtPAL2在花芽发育初期到成熟阶段都表达,而AtPAL1只在花芽发育的成熟阶段表达[4,7]。另外,突变体分析表明,尽管AtPAL1、AtPAL2、AtPAL3、AtPAL4四突变体虽然没有完全丧失PAL活性(约为野生型的10%),但是却极度矮化和不育,特别是在幼苗发育早期,因此,完全破坏PAL基因很可能导致其致死。另外,在AtPAL1、AtPAL2、AtPAL3三突变体和AtPAL1、AtPAL2、AtPAL3、AtPAL4四突变体中没有发现花粉粒,且这些突变体都是不育的,这与之前报道的反义抑制矮牵牛花药中黄酮类合成引起的雄性不育类似[1]。因此,AtPAL1、AtPAL2、AtPAL3三突变体和AtPAL1、AtPAL2、AtPAL3、AtPAL4四突变体中黄酮类含量大量减少是导致其雄性不育的原因。这些结果支持苯丙烷代谢途径对植物生长和发育是必需的[1]。拟南芥AtPAL受光、温度、水、损伤、重金属和营养元素缺乏等非生物因素的单独或协同诱导,并存在时空表达特性。在胁迫条件下,植物对环境压力作出响应的同时完成各种生命活动。在分子水平上,胁迫应答和耐受基因的诱导表达有助于植物适应不利的环境条件[17]。光质和光照都会影响AtPAL的表达。在转录水平上光增加了3周龄拟南芥植株的AtPAL1表达量[4,18],白光处理缓慢积累了AtPAL1的转录,最高是在光照2~4h[4]。虽然与暗培养相比光培养幼苗AtPAL1的表达量有所增加,但它们的表达趋势却一致,且培养3d的幼苗表达量最高,表明无论是光培养还是暗培养,在发育的幼苗中AtPAL1的表达受到严格的时间调控[17]。蓝光处理增加了暗培养白化苗与饱和红光培养幼苗中AtPAL1的转录表达量[17,19],虽然表达趋势一致,但是蓝光处理时培养3d的幼苗中AtPAL1表达量高于培养7d的幼苗[19]。此外,与蓝光相比,紫外光更加促进了AtPAL1的表达[17]。有趣的是,光照强度还能引起对环境条件敏感的AtPAL1、AtPAL2双突变体可育性的变化,因为光照强(120μEm-2S-1)时AtPAL1、AtPAL2双突变体可育,光照弱(50μEm-2S-1)时AtPAL1、AtPAL2双突变体则不育[1]。低温只诱导了叶中AtPAL1和AtPAL2的表达,并且受光和转录水平的调控,这可能与它们在转录起始位点上游330bp内光应答的SORLIP基序相关,因为AtPAL3和AtPAL4缺乏这种基序也不受低温诱导[13,18]。面对不同种类的水分胁迫,AtPAL2的响应机制则不同,Nacl引起水分胁迫时,在叶中的表达量减少,根中的表达量增加;而PEG引起水分胁迫时,AtPAL2的表达则不受任何影响[1]。损伤和氯化汞胁迫处理,瞬时和大量地诱导了AtPAL1的转录表达[4]。缺氮处理时AtPAL1和AtPAL2的转录增加了2倍[10]。非致病菌对AtPAL1的影响远大于致病菌,AtPAL1在非致病型寄生霉菌(Peronosporaparasitica)和非致病型西红柿丁香假单胞菌(Pseudomonassyringaepv.tomato)的非亲和互作中大量表达,而在致病型寄生霉菌(Peronosporaparasitica)和致病型西红柿丁香假单胞菌亲和互作中很少诱导表达[20-21]。叶与西红柿丁香假单胞菌非亲和互作中,2d后在未接种的根中诱导了AtPAL2的表达,推测为遭受细菌侵害时,叶片会通过信号传递来诱导植株获得系统性抗性;而叶与西红柿丁香假单胞菌亲和互作中,在叶中专性地瞬时(1d)诱导了AtPAL2的表达,2d后快速下降到起始水平,表明两种西红柿丁香假单胞菌株通过器官特异性的方式诱导了AtPAL2的表达[7]。这些都反应了面对不同的致病菌,AtPAL采取了不同的响应机制。拟南芥中AtPAL表达与苯丙烷类的合成密切相关,由苯丙烷类合成的木质素不仅与植物的发育和结构适应相关,同时也抵御病原物微生物的侵染;苯丙烷类也是黄酮类、水杨酸、可溶性的苯丙烷类和芥子酸酯等化合物合成的前体,它们在拟南芥生物胁迫和非生物胁迫中发挥了重要作用。PAL是苯丙烷类代谢途径的关键酶,也是合成木质素的关键酶。木质素的形成可以增加细胞壁的厚度,增加组织木质化程度,形成病原菌入侵的机械屏障[6],是植物维管束系统次生加厚细胞壁的主要结构复合物,对维持植物茎直立及传导水、矿物质和光合产物具有重要的作用[1]。在拟南芥生长发育过程中,木质素的合成虽然与AtPAL4相关,但主要与木质化组织中AtPAL1和AtPAL2组成型和协同表达相关[1,4,7],而且AtPAL1或AtPAL2单突变体都没有显著地减少木质素含量,表明AtPAL1和AtPAL2同工酶之间的互作保障了木质素的生物合成,避免了对植物结构造成的不利影响[7,13]。为了防止紫外光造成的损伤,AtPAL1在幼苗中快速表达以积累花色素[4],使植物具备一定的选择优势[17],而且花色素的积累受到严格的发育控制和光受体调节[2]。为了防止病菌感染[4.20-21],AtPAL1以结构适应的方式逐步诱导,参与木质素的生物合成[7],木质素不仅加厚了植物细胞壁使其抵御病菌水解酶的攻击,同时,植物细胞之间通过自由基调节木质素前体的聚合,从而木质化病菌[20]。另外,AtPAL1也参与水杨酸的生物合成,在抑制寄生霉菌时水杨酸的作用远大于木质素抗性作用[20]。低温胁迫时,AtPAL1和AtPAL2协同诱导,使植株可溶性的苯丙烷类、芥子酸酯和黄酮类增加以耐低温[13,21]。此外,低温胁迫时AtPAL1诱导具有光依赖性,与花青素的积累紧密相关,但是在拟南芥耐冻过程中不需要这些色素[18]。总之,AtPAL家族是拟南芥生长发育所必需,它们在植物生长、发育和环境响应时发挥着不同和冗余的双重功能。目前的研究主要集中在AtPAL1和AtPAL2,今后应加强对AtPAL3和AtPAL4的研究。同时,更应该重视拟南芥4个PAL基因家族的不同成员与下游何种代谢支路形成何种代谢通道,以及这种代谢通道在何时、何部位形成何种代谢产物及其发挥的生理功能研究,以便调控PAL基因的表达,有目的地改变下游代谢产物的组成和含量。',)
提供拟南芥苯丙氨酸解氨酶(PAL)基因的研究进展会员下载,编号:1700747745,格式为 docx,文件大小为6页,请使用软件:wps,office word 进行编辑,PPT模板中文字,图片,动画效果均可修改,PPT模板下载后图片无水印,更多精品PPT素材下载尽在某某PPT网。所有作品均是用户自行上传分享并拥有版权或使用权,仅供网友学习交流,未经上传用户书面授权,请勿作他用。若您的权利被侵害,请联系963098962@qq.com进行删除处理。

 下载
下载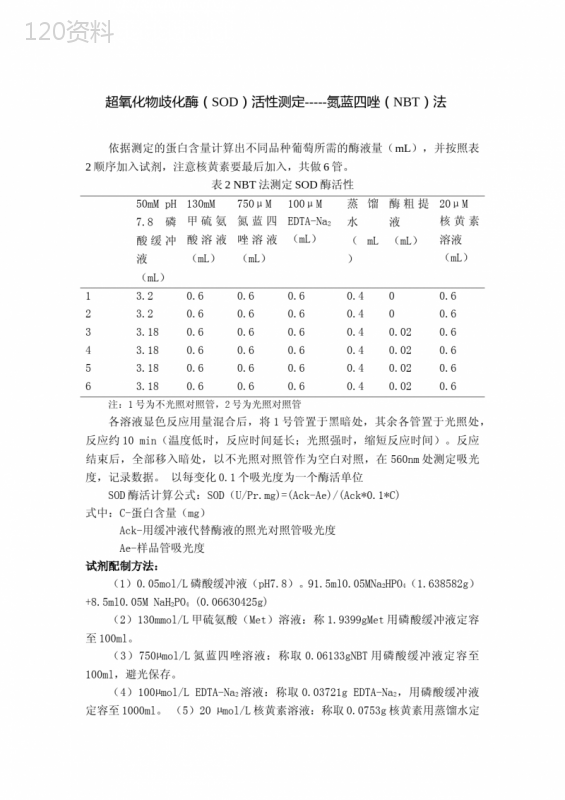 下载
下载 下载
下载 下载
下载 下载
下载 下载
下载 下载
下载 下载
下载